Effects of dietary energy levels on physiological parameters and reproductive performance of gestating sows over three consecutive parities
Article information

Abstract
Objective
This experiment was to evaluate the effects of the dietary energy levels on the physiological parameters and reproductive performance during gestation over three parities in sows.
Methods
A total of 52 F1 gilts (Yorkshire×Landrace) were allotted to one of four dietary treatments using a completely randomized design. The treatments contained 3,100, 3,200, 3,300, or 3,400 kcal of metabolizable energy (ME)/kg diet but feed was provided at 2.0, 2.2, and 2.4 kg/d in the first, second and third parity, respectively.
Results
The body weight and body weight gain during gestation increased as the dietary energy level increased (p<0.05, and p<0.01) in the first parity. In the second parity, the body weight of sows was the lowest (p<0.05) when 3,100 kcal of ME/kg treatment diet was provided. The body weight was higher as the dietary energy level increased (p<0.05) during the gestation period in the third parity. During lactation, the voluntary feed intake of lactating sows tended to decrease when gilts were fed higher energy treatment diet (p = 0.08) and the body weight, body weight gain were increased by dietary energy level during gestation (p< 0.05). Backfat thickness was not affected by dietary treatment during the gestation period in three parities, interestingly backfat change from breeding to d 110 of gestation was higher as the dietary energy level increased at the first parity (p<0.05). When gilts were fed 3,400 kcal of ME/kg treatment diet a higher number of weaning piglets was observed in the first parity (p<0.05). The highest culling rate (69%) was seen when gestating sows were fed 3,100 kcal/kg ME treatment diet during three parities.
Conclusion
In conclusion, the adequate energy intake of gestating sows should be 6,400 or 6,600 kcal of ME/d, 7,040 or 7,260 kcal of ME/d, and 7,680 or 7,920 kcal of ME/d for parity 1, 2, and 3, respectively.
INTRODUCTION
Gestation diets for gilts and sows are of central importance to the swine industry because of their importance to reproductive productivity and longevity of the animal. Jang et al [1] also indicated that the energy intake during gestation should be limited to control body weight gain and maintain an appropriate body condition, especially, in sows from the first to third parity, as adequate energy consumption is required during gestation for the maintenance of body maturation, the growth of the fetus and body preservation. With the development of the genetic potential, many studies were performed to evaluate the nutrient requirement for modern sows. Long et al [2] stated that the provision of high energy feed during gestation caused increased body weight and a backfat thickness loss during lactation. Also, the model developed by NRC [3] suggested that the energy requirement of the gestating gilt and sow should be between 6,678 and 8,182 kcal of metabolizable energy (ME)/d. However, data on the development of the energy level during gestation and its effect on successive parities of gestating sows are lacking. Therefore, the objective of the study was to evaluate the optimum dietary energy level that produced the best physiological parameters and reproductive performance in high-producing modern sows over three consecutive parities.
MATERIALS AND METHODS
The protocol for the present experiment was approved by the Seoul National University Institutional Animal Care and Use Committee (SNUIACUC; SNU-160819-9) in Republic of Korea.
Animal
A total of 52 gilts (Yorkshire×Landrace) weighing approximately 85 kg were selected and housed in an 11×14 m barn. The sows were provided feed and water ad libitum until 120 kg of body weight was reached and were then moved to an individual gestation stall cage with a concrete slatted floors (0.64× 2.40 m). The sows were fed 800 g of an individual diet, twice daily for an average daily gain of 750 g/d. Gilts were mated at an average body weight of 135.82±0.85 kg after three or four estrus cycles. Estrus was diagnosed twice daily in the presence of a mature boar, using the backfat pressure test. Gilts and weaning sows were artificially inseminated with fresh diluted semen (Darby A.I. center, Chungju, Korea) twice at a 12 h interval. A total of 52 crossbred gilts (Yorkshire×Landrace) with 135.82±0.85 kg body weight (BW) were allotted to 4 dietary treatments by BW and backfat thickness in a completely randomized design with 13 replicates. Pregnancy of gilts and sows were diagnosed by an ultrasound analyzer (Easyscan, Dong-jin BLS Co., Ltd., Gwangju, Gyeonggi, Korea) on days 30 and 60 after mating.
Experimental design and animal management
Experimental diets and treatment of sows were not changed in the whole experiment period. Experimental diets for gestating gilts and sows were formulated to contain 13.08% crude protein (CP), 0.86% lysine, 0.90% calcium, and 0.70% phosphorus, with an energy content of 3,100, 3,200, 3,300, or 3,400 kcal of ME/kg and diets were provided daily at 2.0 kg/d for the 1st parity, 2.2 kg/d for the 2nd parity, 2.4 kg/d for the 3rd parity and 3 kg from weaning to estrus. Lactation diets contained 3,265 kcal ME/kg, 17.07% CP, 1.26% lysine, 0.90% calcium, and 0.70% phosphorus (Table 1). All other nutrients were formulated to meet or exceed the NRC requirements [3]. Gilts and sows were housed in temperature-controlled rooms and placed in an individual crate (2.4× 0.65 m2) with a concrete floor until 110 d of gestation. After 110 d of gestation, pregnant gilts and sows were washed and moved into farrowing crates (2.4×1.8 m2). During lactation, all sows were fed the same commercial lactation diet. After farrowing, the lactation diet was increased gradually from 1.0 kg/d until 5 d postpartum and then provided ad libitum during lactation. Weaning was at approximately 21 d and sows returned to stall cage again for the next reproductive cycle. Gilts and sows were excluded from the feeding trial for reproductive problems and lameness.

Formula and chemical composition of gestation and lactation diets (as dry matter basis)
Measurements and analysis
The BW and backfat thickness at the P2 position of the sows were measured. Blood samples were collected at breeding, 110 days of gestation, 24 h post-farrowing and 21 days of lactation from sows. The number of total piglets born, piglets born alive, still born, and mummified fetuses as well as the piglet BW were recorded. The fat and protein mass of primiparous and multiparous sows were calculated using the equations of Dourmad et al [4].
Blood samples were collected from the jugular vein of sows with tubes (serum and EDTA tube, BD Vacutainer, Berkshire, UK) and centrifuged immediately at 3,000 rpm at 4°C, and then, samples were stored at −20°C until later analysis. Colostrum and milk were collected from the first and second teats at 24 h and 21 d postpartum after an intravascular injection of 5 IU oxytocin (Komi oxytocin inj. Komipharm International Co., Ltd., Siheung, Korea) in the ear. All samples were stored at −20°C until analysis. A proximate analysis of colostrum and milk samples was conducted using a Milkoscan FT 120 (FOSS Electric, Sungnam, Korea). The glucose and blood urea nitrogen (BUN) concentrations were analyzed using a kinetic UV assay (Glucose Hexokinase Kit; UREA/BUN Kit, Roche, Mannheim, Germany). Plasma free fatty acid (FFA) concentrations were determined according to the colorimetric Acyl-CoA synthetase Acyl-CoA oxidase method [5] using a commercial kit (Wako FFA c Kit; Wako chemical, Osaka, Japan). The fatty acid content in colostrum was analyzed on an Agilent 7890 gas liquid chromatograph (Agilent Technologies, Palo Alto, CA, USA) equipped with a flame ionization detector and an SP-2560 (i.d. 100 m×0.25 mm×0.20 μm) film column. Nitrogen was used as carrier gas, injector core temperature was 250°C, detector temperature was 260°C and column temperature was programmed to begin at 170°C and then increase to 250°C and remain at 240°C for 40 min. Chromatography was calibrated with a mixture of 37 different fatty acids (FAME 37; Supelco Inc., Bellefonte, PA, USA) and this standard containing fatty acids ranging from C4:0 to C24:1n9 and samples were added 250 μL of internal standard spike solution (Pentadecanoic acid; Sigma-Aldrich, Darmstadt, Germany) by the method of AOAC [6].
Statistical analysis
Data were analyzed by analysis of variance with a completely randomized design using the general linear model procedure implemented in SAS. The least squares means were calculated for each independent variable. Orthogonal polynomial contrasts were used to determine the linear and quadratic effects by increasing the dietary energy level during gestation for all measurements of sows and piglets. The individual sows and their litters were used as the experimental unit. The alpha level used for the determination of significance for all analyses was 0.05 and for the determination of trends was p>0.05 and p<0.10
RESULTS
The BW and body weight gain during gestation increased as the dietary energy level increased (linear, p<0.05, and p<0.01, respectively, Table 2) in the first parity. In the second parity, BW was the lowest (quadratic, p<0.05) in the 3,100 kcal/kg ME treatment with a higher body weight gain (linear, p = 0.07, Table 2). The BW increased with an increasing energy level (linear, p<0.05) during gestation in the third parity (Table 2). During lactation, an increasing energy level led to lower BW, body weight gain and overall body weight gain (linear, p<0.05, p = 0.06, respectively) in the first parity (Table 2).

Effects of dietary energy level on the body weight of gestating and lactating sows over three consecutive parities
Back fat thickness was not affected by the diet during gestation in parity 1, 2, or 3. However, back fat difference from breeding to d 110 of gestation increased linearly (p<0.05) as the dietary energy level increased in parity 1 (Table 3).

Effects of dietary energy level on the back-fat of gestating and lactating sows over three consecutive parities
The estimated fat and protein masses were calculated based on BW and backfat thickness [5]. The fat mass and protein mass were higher as the energy level increased (linear, p<0.01, and p< 0.05, respectively) during gestation in parity 1 (Table 4). During lactation, the fat mass and protein mass decreased (linear and quadratic, p<0.01, and p<0.05, respectively) with an increased dietary energy level in parity 1 (Table 5).

Effects of the dietary energy level on the estimated fat and protein mass of gestating sows over three consecutive parities

Effects of energy level on the estimated fat and protein mass of lactating sows over three consecutive parities
The voluntary feed intake of sows tended to decrease (linear, p = 0.08) when the dietary energy level increased in parity 1 (Table 6). The weaning to estrus interval (WEI) was not significantly affected by treatment over the three parities (Table 6). The culling rate was the highest in the 3,100 kcal/kg ME treatments.

Effects of dietary energy level on the lactation feed intake, weaning to estrus interval after lactation and culling rate of sows over three consecutive parities
The 3,400 kcal/kg ME treatment showed the highest number of weaning pigs per litter (quadratic, p<0.05) in the first parity (Table 7).

Effects of dietary energy level on the reproductive performance of sows over three consecutive parities
The BUN concentration in sows tended to be increased by with the dietary energy level at d110 of gestation and 24 h postpartum (linear, p = 0.06, and p = 0.07, respectively) in parity 1 (Figure 1). The glucose concentration was higher (linear, p = 0.07) and tended to be the lowest in 3,300 kcal/kg ME (quadratic, p = 0.06) treatment at 24 h postpartum in parities 1, and 3 (Figure 2). Similarly, the 3,300 kcal/kg ME treatment had a lower glucose concentration at d 110 in the third parity (quadratic, p = 0.06, Figure 2). The FFA concentration was not affected by dietary energy level (Figure 3).
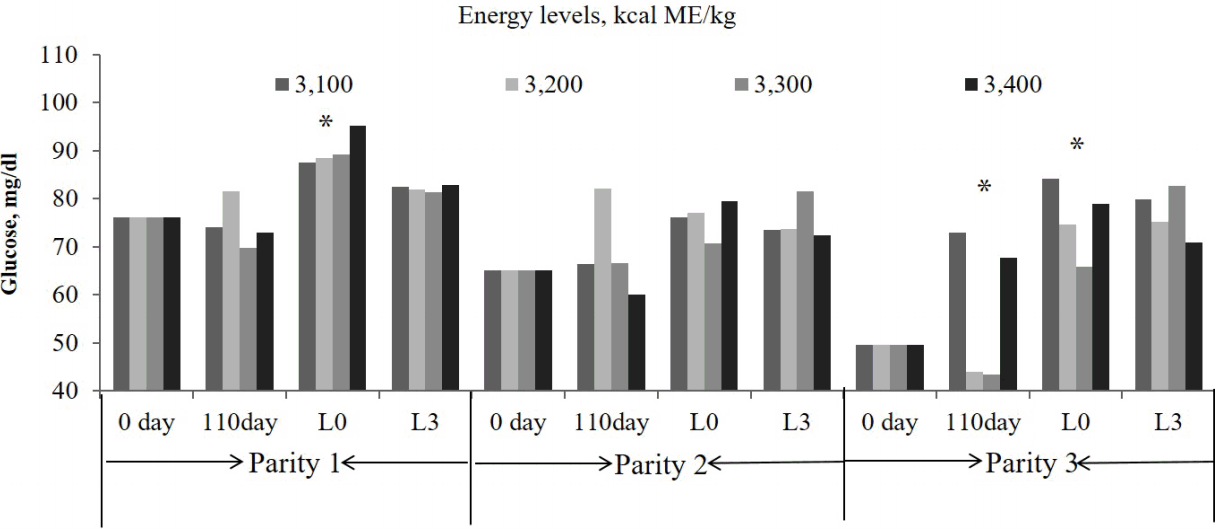
Effects of the dietary energy level on the blood urea nitrogen concentration in the blood of sows over three consecutive parities (* p<0.1).
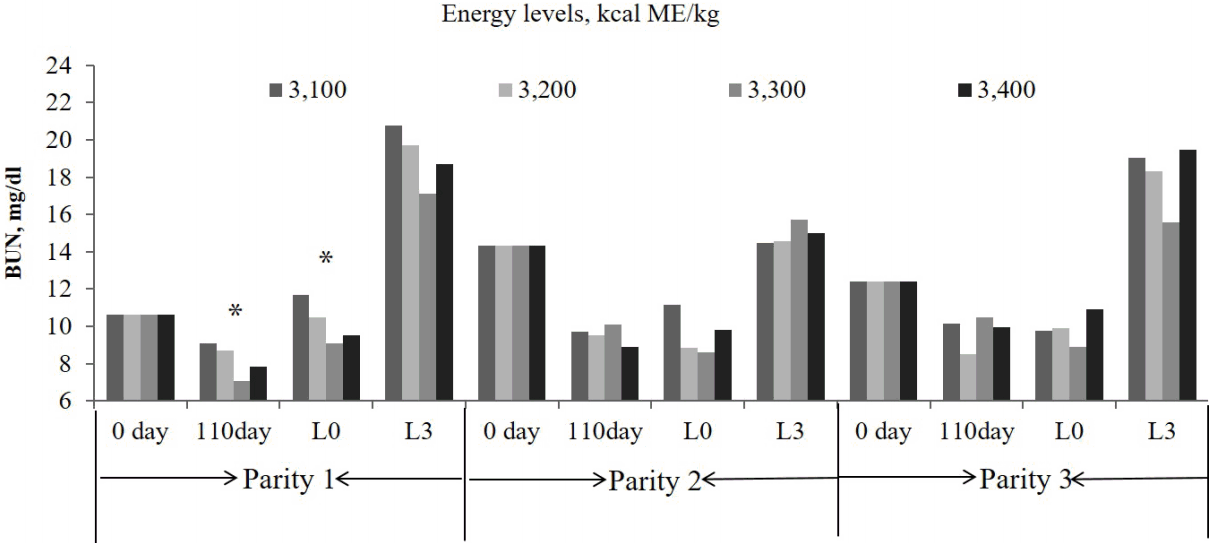
Effects of the dietary energy level on the glucose concentration in the blood of sows over three consecutive parities (* p<0.1).
Effects of the dietary energy level on the free fatty acid concentration in the blood of sows over three consecutive parities
No statistical differences were evident in the fat content of the colostrum and milk (Table 9). The colostrum fatty acids were also not affected by dietary energy level (Table 10).

Effects of dietary energy level in gestating sows on the fat content in colostrum and milk of lactating sows over three consecutive parities

Effects of dietary energy level in gestating sows on the fatty acids compositions of colostrum in primiparous lactating sows over three consecutive parities (mg/g)
DISCUSSION
During gestation, the maternal BW of sows should gain 25 kg per parity over three or four parities [7]. During the gestation period there is a total 45 kg of weight gain by the sow; 20 kg is the weight of the placental and other products of conception out of the total weight [8]. In this study, all treatments showed a 45 kg of body weight gain during gestation except the 3,100 kcal/kg ME treatment in the first parity. This result demonstrated that an energy level of 3,100 kcal/kg ME might not be high enough to increase the BW during gestation. In backfat thickness, all treatments produced a backfat thickness greater than 20 mm backfat at farrowing and 16 mm at weaning. Averette Gatlin et al [9] suggested that the effect of the energy level during gestation on BW and body weight gain is highly related to the BW, which may be attributed to a higher backfat thickness due to a higher energy level [2]. However, Young et al [10] indicated that higher energy intake during gestation reduced the voluntary feed intake during lactation. Our results suggested that BW and backfat loss increased with dietary energy level in the first parity, which was in agreement with previous studies, and demonstrated that the provision of high energy feed during gestation caused increased BW and a loss of backfat thickness during lactation [2].
Fat tissue and protein tissue were increased during gestation, whereas fat and protein mass decreased with increasing energy level during lactation in the first parity, indicating that the energy supply was important factor to maintain adequate BW and back fat thickness for subsequent reproductive cycles in sows. These results were in agreement with previous studies, which demonstrated that N retention was increased by a high energy level [11] and higher feed intake during gestation [12].
Previous studies suggested that unbalanced nutrient intake caused several common reproductive problems, such as an increase in the interval from weaning to estrus [13], an increased incidence of anestrus after weaning, and a decreased conception rate [14]. However, in this study, WEI was not affected by treatment, and the 3,100 kcal/kg ME treatment had the highest culling rate (68%) because of pregnancy failure and anestrus after weaning, which was in agreement with Kongsted [15], who suggested that a low energy intake during gestation might increase the risk of culling. It is well documented that a late WEI is related to a high glucose and low FFA concentration in weaned sows [16]. In this study, the plasma glucose and FFA concentration of sows at weaning was not affected by treatment, indicating that the dietary energy level did not affect the WEI.
Prunier et al [17] suggested that there was no treatment effect on number of embryos when the energy intake was increased from moderate (28 MJ d/ME) to high (37 MJ d/ME). Also, previous studies suggested that a high energy supply (50.1 and 48.6 MJ d/ME) 3 days after mating or immediately after mating did not affect the number of embryos or the litter size in sows [18,19]. Similar results were also observed in this study.
It is well documented that increased energy intake during late gestation can positively affect fetal growth [3]. However, no effect was observed on litter birth weight and individual piglet birth weight (Table 8), which was consistent with results of Long et al [2], who demonstrated that the average piglet BW at farrowing was not affected by different energy levels in the gestation diet. Similarly, Piao et al [20] also suggested that an increased feed intake during gestation did not increase litter weight or individual piglet weight. In this study, the 3,400 kcal ME/kg treatment showed the highest weaning litter size in the first parity. However, feed intake decreased in the 3,400 kcal ME/kg treatment more than the other treatments with an increased BW, backfat thickness loss, and a decreased culling rate.

Effects of dietary energy level on the progeny growth performance of sows over three consecutive parities
It is very well known that BUN is connected to retention of nitrogen in the body [21]. In this study, the serum BUN concentration tended to decrease with an increasing energy level at 110 days of gestation and 24 hours post-farrowing in the first parity, which was consistent with the results of Ruiz et al [22], who reported that the BUN concentration was lower in swine that were fed a high energy diet compared to pigs fed a low energy diets. These results might suggest that the energy intake in sows affects the protein metabolism during gestation. An increased energy level during gestation could decrease glucose utilization and subsequently feed intake during lactation [23]. Moreover, an increased feed intake in gestating gilts may cause sows to become insensitive to insulin, which presents a smaller response in glucose clearance and decreased feed during lactation [20]. In this study, the glucose concentration was increased with a higher energy level 24 hours post-farrowing in the first parity, but was lowest in the 3,300 kcal/kg ME treatments at 110 days of gestation and 24 hours post- farrowing in the third parity. Therefore, the effect of insulin on feed intake during lactation might depend on the body condition of the sows and the glucose metabolites, and this can explain our results of higher bodyweight and backfat thickness loss with an increased energy level during lactation in parity 1, but not in parity 3.
The chemical composition of the colostrum and milk of sows is variable due to the dietary regimen [24] and the body condition of the sows [25]. Feeding a fatty diet in late gestation increased the total lipids in colostrum [26]. However, Yang et al [27] reported that there was no effect on the colostrum composition when the energy level was increased from 13.7 to 14.2 MJ of ME/kg in the gestation diet. In this study, no significant difference in the fat content of sow colostrum and milk was noted, which is in agreement with Williams et al [28], who demonstrated that the chemical composition of colostrum and milk was not affected by dietary energy level during gestation because the sow mobilized its body reserves to compensate for the nutrient deficiency.
The fatty acid composition of colostrum was affected by the dietary fat level [29] and type of fat provided in the diet [30], which was inconsistent with our results, which showed that the fatty acids composition of colostrum was not influenced by energy level during gestation. However, studies on the effect of energy level during gestation on the fatty acid composition of colostrum are limited, and further studies are still warranted to elucidate a detailed mechanism. Consequently, the adequate energy intake of gestating sows was 6,400 or 6,600 kcal of ME/d for parity 1, 7,040 or 7,260 kcal of ME/d for parity 2, and 7,680 or 7,920 kcal of ME/d for parity 3.
ACKNOWLEDGMENTS
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Agri-Bio Industry Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA)(314022-3).
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.